O Citoplasma e suas Estruturas
O citoplasma tem uma estrutura complexa com uma emaranhada rede
de canalículos, e os espaços que permeiam essa rede são
preenchidos por um material que constitui o hialoplasma ou
citoplasma fundamental.
No espaço entre a membrana plasmática e a nuclear encontram-se:
retículo endoplasmático, mitocôndrias, complexo ou aparelho
golgiense, cloroplastos, centríolos ou centrossomos, ribossomos,
lisossomos, vacúolos e microtúbulos.
O retículo endoplasmático é um sistema de canalículos que
confluem para pequenos vacúolos ou se abrem em bolsas achatadas
ou cisternas, formando uma emaranhada rede que ocupa a maior
parte do citoplasma.
Todo esse sistema é delimitado por membranas lipoprotéicas.
Ele só é encontrado nas células dos eucariontes, estendendo-se
muitas vezes desde a membrana plasmática até a carioteca,
aumentando grandemente a superfície interna celular.
Em alguns pontos observa-se a presença de grande número de
ribossomos aderidos à face externa das membranas do retículo
endoplasmático. Os ribossomos são grânulos formados de proteínas
e RNA que atuam na síntese protéica.
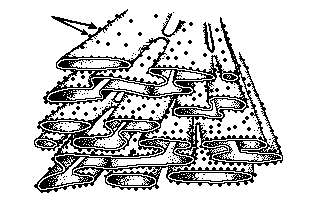
As regiões do retículo
endoplasmático onde se acumulam os ribossomos caracterizam o
retículo endoplasmático granuloso (REG). Nas células
glandulares e naquelas onde a síntese protéica é intensa, o REG
se mostra muito desenvolvido. Quando o retículo endoplasmático
apresenta membranas lisas, sem ribossomos, é chamado de
retículo endoplasmático não-granuloso (RENG), funcionando na
síntese de lipídios e como via de transporte de substâncias de
um ponto para outro do citoplasma.
O RENG desempenha as seguintes funções:
Nele se realizam reações enzimáticas facilitadas por sua ampla
superfície;
Transporta substâncias através da formação de vesículas;
Armazena substâncias por meio de vacúolos;
Sintetiza lipídios como o colesterol, a lecitina e os hormônios
sexuais.
O REG, além dessas funções, sintetiza proteínas graças à
presença dos ribossomos.
O complexo golgiense é uma região especial do RENG que se
mostra como um empilhamento de bolsas achatadas ou cisternas,
rodeadas de pequenos vacúolos ou vesículas que se desprendem da
bolsa por brotamento. Existe em todas as células eucariotas.
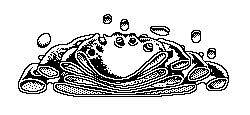
Desenvolve as seguintes atividades:
Acúmulo de proteínas sintetizadas no REG para posterior
eliminação por clasmocitose;
Produção de glicoproteínas graças à associação de moléculas de
proteínas provenientes do REG com polissacarídeos sintetizados
no próprio complexo golgiense pela polimerização de
monossacarídeos obtidos pela alimentação;
Síntese de esteróides, como sucede em células das glândulas
supra-renais e nas gônadas.
As mitocôndrias são encontradas em todas as células
eucariotas. O seu contorno é delimitado por uma dupla membrana
lipoprotéica. A externa é lisa e a interna é pregueada ou
franjada, com numerosas dobras perpendiculares ao eixo do
orgânulo chamadas de cristas mitocondriais. Essas cristas
dividem o interior da mitocôndria em lojas que fazem
continuidade entre si. O interior do orgânulo é preenchido por
um material homogêneo, a matriz mitocondrial.
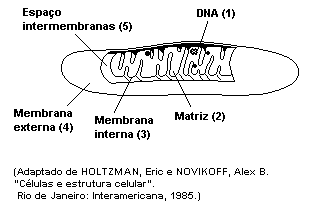
Desenvolvem importante atividade nos processos metabólicos
celulares.
Reprocessam a energia contida nas moléculas dos compostos
orgânicos obtidos pela alimentação (respiração celular),
transferindo o acúmulo energético para outras moléculas
especializadas para armazenamento e liberação rápida de energia.
Elas produzem moléculas de ATP (adenosina trifosfato).
Dispõem de moléculas de DNA e RNA, ribossomos, sintetizam suas
próprias proteínas e se auto-reproduzem.
Plastos ou plastídios
são orgânulos de estrutura membranosa encontrados em todas as
células vegetais e em alguns protistas como as euglenófitas e
diatomáceas. São inexistentes nas moneras, nos fungos e nos
animais.
Os plastos são dotados de uma dupla membrana de natureza
lipoprotéica. A externa é lisa e a interna faz dobras ou franjas
que se dispõem como lâminas paralelas no sentido do maior eixo
do plasto. Essas dobras se chamam lamelas e na sua estrutura
química se encontra a clorofila, substância extremamente
importante para a realização da fotossíntese. São portadores de
DNA, RNA, ribossomos e se auto-reproduzem.
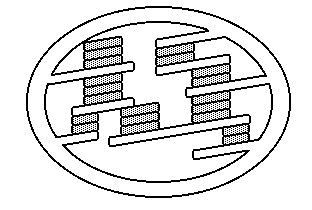
Compreendem algumas variedades que se distinguem essencialmente
pela cor, em função dos pigmentos de que são portadores:
Cloroplastos (verdes – com clorofila);
Leucoplastos (brancos – com amido ou lipídios);
Cromoplastos ou cromatóforos (amarelos – xantoplastos; pardos –
feoplastos; vermelhos – eritroplastos).
De qualquer cor, todos eles são cloroplastos disfarçados, que
acumularam pigmentos diversos, encobrindo a clorofila. Os
leucoplastos são brancos pelo acúmulo de amido, lipídios ou
proteínas. São abundantes nas células de armazenamento das
raízes, caules e frutos. Os cromoplastos acumulam pigmentos
carotenóides que lhes dão a cor amarela, alaranjada ou vermelha.
Podem ser vistos nas folhas, nos frutos, na cenoura, beterraba,
etc.
Os lisossomos são minúsculas vesículas delimitadas por
membrana lipoprotéica e espalhadas pelo citoplasma. Existem
sempre nas células animais. Desempenham papel importante na
realização da digestão intracelular, pois encerram no seu
interior razoável quantidade de enzimas hidrolisantes
(proteolíticas, lipolíticas e glicolíticas).
Atuam intensamente na autólise e autofagia.
Quando um orgânulo envelhecido é digerido no lisossomo ocorre a
autofagia e quando há necessidade da destruição total da célula
os lisossomos se rompem e seu conteúdo se derrama no citoplasma,
realizando a autólise (metamorfose dos sapos, por exemplo).
O desenvolvimento de seres multicelulares depende da morte
programada de certas células. Esse fenômeno biológico, regulado
por genes, é conhecido como apoptose:
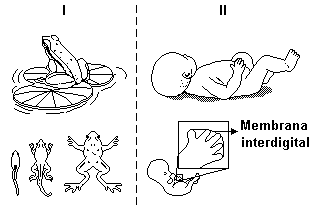
Durante a metamorfose dos anfíbios, desaparecem as guelras, as
nadadeiras e a cauda.
No embrião humano, os sulcos dos dedos das mãos são formados
como conseqüência da morte das células das membranas
interdigitais. A apoptose resulta da ação de enzimas digestivas
presentes nos lisossomos.
A ocorrência de alterações nos genes responsáveis pela apoptose
pode ser transmitida aos descendentes
Os peroxissomos são pequeninas vesículas contendo enzimas
oxidantes e limitadas por membrana lipoprotéica. Originam-se a
partir do RENG, onde acumulam enzimas provenientes do REG.
Dentre as enzimas acumuladas, a de ação mais notável é a
catalase, que oxida a água oxigenada ou peróxido de hidrogênio
(H2O2), decompondo-a em água comum e
oxigênio nascente. A água oxigenada se forma nas células como
produto final de certas reações e tem efeito altamente lesivo.
Os vacúolos dividem-se em três tipos essenciais:
digestivos, pulsáteis e de suco celular.
Os vacúolos digestivos são resultantes da fusão de fagossomos ou
pinossomos com os lisossomos (fagocitose e pinocitose).
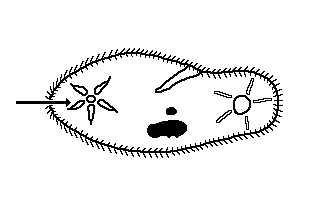
Os vacúolos pulsáteis ou contráteis são encontrados nos
protozoários e euglenófitas, nos quais contribuem para a
manutenção do equilíbrio homeostático, pois eliminam o excesso
de água e alguns derivados nitrogenados do metabolismo protéico,
como amônia.
Os vacúolos de suco celular são observados nas células vegetais.
Contêm água com algumas substâncias e pigmentos. Pequenos e
numerosos nas células vegetais jovens, crescem e se fundem na
célula vegetal adulta, formando poucos e enormes vacúolos
repletos de líquidos que deslocam o citoplasma para a periferia.
O centrossomo, centro-celular ou centríolo é uma
estrutura não membranosa do citoplasma, existente em células de
animais e de vegetais inferiores (algas e briófitas). Só é bem
visível durante a mitose (divisão celular), quando dele se
irradiam os microtúbulos que orientam a distribuição dos
cromossomos para as células-filha. São importantes também na
formação de flagelos e cílios.
Os ribossomos são grãos formados por RNA e proteínas.
Estão presentes em todos os seres vivos, até nos mais simples
como os PPLO. Podem ser encontrados dispersos no citoplasma
(procariontes) ou ligados ao REG (eucariontes).
Leia mais...




